Можно сказать, что молекулярная биология исследует проявления жизни на неживых структурах или системах с элементарными признаками жизнедеятельности (которыми могут быть отдельные биологические макромолекулы, их комплексы или органеллы), изучая, каким образом ключевые процессы, характеризующие живую материю, реализуются посредством химических взаимодействий и превращений.
Выделение молекулярной биологии из биохимии в самостоятельную область науки продиктовано тем, что её главной задачей является изучение структуры и свойств биологических макромолекул, участвующих в различных процессах, выяснение механизмов их взаимодействия. Биохимия же занимается изучением собственно процессов жизнедеятельности, закономерностей их протекания в живом организме и превращений молекул, сопровождающих эти процессы. В конечном счёте, молекулярная биология пытается ответить на вопрос, зачем происходит тот или иной процесс, тогда как биохимия отвечает на вопросы где и как с точки зрения химии происходит рассматриваемый процесс.
История
Молекулярная биология как отдельное направление биохимии начала формироваться в 30-х годах прошлого столетия. Именно тогда для углублённого понимания феномена жизни возникла необходимость в целенаправленных исследованиях на молекулярном уровне процессов хранения и передачи наследственной информации в живых организмах. Тогда и определилась задача молекулярной биологии в изучении структуры, свойств и взаимодействия нуклеиновых кислот и белков. Термин «молекулярная биология» был впервые употреблен английским учёным Уильямом Астбери в контексте исследований, касавшихся выяснения зависимостей между молекулярной структурой и физическими и биологическими свойствами фибриллярных белков, таких, как коллаген, фибрин крови или сократительные белки мышц.
На заре возникновения молекулярной биологии РНК считалась компонентом растений и грибов, а ДНК рассматривалась как типичный компонент животных клеток. Первым исследователем, доказавшим, что ДНК содержится в растениях, был Андрей Николаевич Белозерский , выделивший в 1935 году ДНК гороха. Это открытие установило тот факт, что ДНК является универсальныой нуклеиновой кислотой, присутствующей в клетках растений и животных.
Серьёзным достижением стало установление Джорджем Бидлом и Эдуардом Татумом прямой причинно-следственной связи между генами и белками. В своих экспериментах они подвергали клетки нейроспоры (Neurospora crassa ) ретгеновскому облучению, вызывавшему мутации. Полученные результаты показали, что это приводило к изменению свойств специфических ферментов.
В 1940 году Альбер Клод выделил из цитоплазмы животных клеток цитоплазматические РНК-содержащие гранулы, которые были меньше митохондрий. Он назвал их микросомами. Впоследствии при исследовании структруы и свойств выделенных частиц была установлена их основополагающая роль в процессе биосинтеза белка. В 1958 году на первом симпозиуме, посвящённом этим частицам, было принято решение называть эти частицы рибосомами.
Ещё одним важным шагом в развитии молекулярной биологии стали опубликованные в 1944 г. данные эксперимента Освальда Эвери, Колина МакЛауда и Маклина МакКарти, показавшие, что причиной трансформации бактерий является ДНК. Это было первое экспериментальное доказательство роли ДНК в передаче наследственной информации, развенчавшее бытовавшее ранее представление о белковой природе генов.
В начале 50-х годов Фредерик Сэнгер показал, что белковая цепь является уникальной последовательностью аминокислотных остатков. В конце 50-х годов Макс Перуц и Джон Кендрю расшифровали пространственное строение первых белков. Уже в 2000 году были известны сотни тысяч природных аминокислотных последовательностей и тысячи пространственных структур белков.
Примерно в то же время исследования Эрвина Чаргаффа позволили ему сформулировать правила, описывающие соотношение азотистых оснований в ДНК (правила гласят, что независимо от видовых различий в ДНК количество гуанина равно количеству цитозина, а количество аденина равно количеству темина), что помогло в дальнейшем сделать величайший прорыв в молекулярной биологии и одно из величайших открытий в биологии вообще.
Это событие произошло в 1953 году, когда Джеймс Уотсон и Фрэнсис Крик , основываясь на работах Розалинды Франклин и Мориса Уилкинса по рентгено-структурному анализу ДНК, установили двухспиральную структуру молекулы ДНК. Это открытие позволило ответить на принципиальный вопрос о способности носителя наследственной информации к самовоспроизведению и понять механизм передачи такой информации. Этими же учеными был сформулирован принцип комплементарности азотистых оснований, имеющий ключевое значение для понимания механизма образования надмолекулярных структур. Это принцип, применяемый теперь для описания всех молекулярных комплексов, позволяет описывать и предсказывать условия возникновения слабых (невалентных) межмолекулярных взаимодействий, обуславливающих возможность формирования вторичной, третичной и т.д. структуры макромолекул, протекания самосборки надмолекулярных биологических систем, определяющих столь большое разнообразие молекулярных структур и их функциональных наборов. Тогда же, в 1953 году возник научный журнал Journal of Molecular Biology. Его возглавил Джон Кендрю, сферой научных интересов которого было исследование структуры глобулярных белков (Нобелевская премия 1962 года совместно с Максом Перуцем). Аналогичный русскоязычный журнал под названием «Молекулярная биология» был основан в СССР В. А. Энгельгардтом в 1966 году.
В 1958 году Фрэнсис Крик сформулировал т.н. центральнаю догму молекулярной биологии: представление о необратимости потока генетической информации от ДНК через РНК к белкам по схеме ДНК→ДНК (репликация, создание копии ДНК), ДНК→РНК (транскрипция , копирование генов), РНК→ белок (трансляция, декодирование информации о структуре белков). Эта догма в 1970 году была несколько поправлена с учётом накопленных знаний, поскольку было открыто явление обратной транскрипции независимо Ховардом Темином и Дэвидом Балтимором: был обнаружен фермент - ревертаза, отвечающий за осуществление обратной транскрипции - образования двуцепочечной ДНК на матрице одноцепочечной РНК, которое происходит у онкогенных вирусов. Следует отметить, что строгая необходимость потока генетической информации от нуклеиновых кислот к белкам до сих пор остаётся основой молекулярной биологии.
В 1957 году Александр Сергеевич Спирин совместно с Андреем Николаевичем Белозерским показали, что, при существенных различиях в нуклеотидном составе ДНК из разных организмов, состав суммарных РНК сходен. На основании этих данных они пришли к сенсационному заключению о том, что суммарная РНК клетки не может выступать в качестве переносчика генетической информации от ДНК к белкам, поскольку не соответствует ей по своему составу. Вместе с тем они заметили, что существует минорная фракция РНК, которая полностью соответствует по своему нуклеотидному составу ДНК и которая может быть истинным переносчиком генетической инфрмации от ДНК к белкам. В результате они предсказали существование относительно небольших молекул РНК, являющихся по строению аналогами отдельных участков ДНК и выполняющих роль посредников при передаче генетической информации, содержащейся в ДНК, в рибосому, где с использованием этой информации осуществляется синтез белковых молекул. В 1961 году (С. Бреннер , Ф. Жакоб , М. Месельсон с одной стороны и Ф. Гро, Франсуа Жакоб и Жак Моно первыми получили опытное подтверждение существования таких молекул - информационной (матричной) РНК. Тогда же они разработали концепцию и модель функциональной единицы ДНК - оперона, которая позволила объяснить, как именно осуществляется регуляция экспрессии генов у прокариот. Исследование механизмов биосинтеза белка и принципов структурной организации и работы молекулярных машин - рибосом - позволило сформулировать постулат, описывающий движение генетической информации, называемый центральной догмой молекулярной биологии: ДНК - иРНК - белок.
В 1961 году и в течение последующих нескольких лет Хайнрихом Маттэем и Маршаллом Ниренбергом, а затем Харом Кораной и Робертом Холли были проведены несколько работ по расшифровке генетического кода, в результате которых была установлена непосредственная взаимосвязь между структурой ДНК и синтезируемыми белками и определена последовательность нуклеотидов, определяющая набор аминокислот в белке. Также были получены данные об универсальности генетического кода. Открытия были отмечены нобелевской премией 1968 года.
Для развития современных представлений о функциях РНК решающим было открытие некодирующих РНК, сделанное по результатам работ Александра Сергеевича Спирина совместно с Андреем Николаевичем Белозерским 1958 года, Чарльзом Бреннером с соавторами и Солом Шпигельманом 1961 года. Этот вид РНК составляют основную часть клеточных РНК. К некодирующим в первую очередь относятся рибосомные РНК.
Серьёзное развитие получили способы культивирования и гибридизации животных клеток. В 1963 году Франсуа Жакобом и Сиднеема Бреннером были сформулированы представления о репликоне - последовательности неотъемлемо реплицирующихся генов, объясняющей важные аспекты регуляции репликации генов.
В 1967 году в лаборатории А. С. Спирина было впервые продемонстрировано, что форма компактно свёрнутой РНК определяет морфологию рибосомной частицы.
В 1968 году было сделано значительное фундаментальное открытие. Оказаки, обнаружив фрагменты ДНК отстающей цепи при исследовании процесса репликации, названные в честь неё фрагментами Оказаки, уточнила механизм репликации ДНК.
В 1970 году независимо Ховардом Темином и Дэвидом Балтимором было сделано значительное открытие: был обнаружен фермент - ревертаза, отвечающий за осуществление обратной транскрипции - образования двуцепочечной ДНК на матрице одноцепочечной РНК, которое происходит у онкогенных вирусов, содержащих РНК.
Ещё одним важным достижением молекулярной биологии стало объяснение механизма мутаций на молекулярном уровне. В результате серии исследований были установлены основные типы мутаций: дупликации, инверсии, делеции, транслокации и транспозиции. Это дало возможность рассматривать эволюционные изменения с точки зрения генных процессов, позволило разработать теорию молекулярных часов, которая применяется в филогении.
К началу 70-х годов были сформулированы основные принципы функционирования нуклеиновых кислот и белков в живом организме. Было установлено, что белки и нуклеиновые кислоты в организме синтезируются по матричному механизму, молекула-матрица несёт в себе зашифрованную информацию о последовательности аминокислот (в белке) или нуклеотидов (в нуклеиновой кислоте). При репликации (удвоении ДНК) или транскрипции (синтезе иРНК) такой матрицей служит ДНК, при трансляции (синтезе белка) или обратной транскрипции - иРНК.
Таким образом, были созданы теоретические предпосылки для развития прикладных направлений молекулярной биологии, в частности, генетической инженерии . В 1972 году Пол Берг , Герберт Боер и Стэнли Коэн разработали технологию молекулярного клонирования. Тогда ими впервые была получена в пробирке рекомбинантная ДНК. Эти выдающиеся эксперименты заложили основы генетической инженерии, а этот год считается датой рождения этого научного направления.
В 1977 году Фредерик Сэнгер, и независимо Аллан Максам и Уолтер Гилберт разработали различные методы определения первичной структуры (секвенирования) ДНК. Метод Сэнгера, так называемый метод обрыва цепи, является основой современного метода секвенирования. Принцип секвенирования основан на использовании меченых оснований, выступающих в качестве терминаторов в циклической реакции секвенирования. Этот метод получил широкое распространение благодаря возможности быстро проводить анализ.
1976 г. - Фредерик. Сэнгер расшифровал нуклеотидную последовательность ДНК фага φΧ174 длиной 5375 нуклеотидных пар.
1981 г. - серповидноклеточная анемия становится первой генетической болезнью, диагностируемой с помощью анализа ДНК.
1982-1983 открытие каталитической функции РНК в американских лабораториях Т. Чека и С. Олтмана изменило существовавшее представления об исключительной роли белков. По аналогии с каталитическими белками - энзимами, каталитические РНК были названы рибозимами.
1987 год Кери Мюллез открыл полимеразную цепную реакцию, благодаря которой возможно искусственно значительно увеличить количество молекул ДНК в растворе для дальнейшей работы. На сегодняшний день это один из наиболее важных методов молекулярной биологии, применяющийся при исследовании наследственных и вирусных заболеваний, при изучении генов и при генетическом установлении личности и установлении родства и т.п.
В 1990 году одновременно тремя группами учёных был опубликован метод, позволявший быстро получать в лаборатории синтетические функционально активные РНК (искусственные рибозимы или молекулы, взаимодействующие с различными лигандами - аптамеры). Этот метод получил название «эволюция в пробирке». А вскоре после этого, в 1991-1993 года в лаборатории А.Б. Четверина была экспериментально показана возможность существования, роста и амплификации молекул РНК в форме колоний на твёрдых средах.
В 1998 году практически одновременно Крейг Мелло и Эндрю Фаер описали наблюдавшийся ранее при генных экспериментах с бактериями и цветами механизм РНК-интерференции , при котором небольшая двухцепочечная молекула РНК приводит к специфичному подавлению экспрессии гена.
Открытие механизма РНК-интерференции имеет очень важное практическое значение для современной молекулярной биологии. Это явление широко используется в научных экспериментах в качестве инструмента для «выключения», то есть, подавления экспрессии отдельных генов. Особый интерес вызван тем, что этот способ позволяет осуществлять обратимое (временное) подавление активности изучаемых генов. Ведутся исследования возможности применения этого явления для лечения вирусных, опухолевых, дегенеративных и метаболических заболеваний. Следует отметить, что в 2002 году были открыты мутанты вирусы полиомиелита, способные избегать РНК-интерференции, поэтому требуется ещё кропотливая работа для разработки действительно эффективных методов лечения на основе этого явления.
В 1999-2001 годах несколькими группами исследователей определена с разрешением от 5,5 до 2,4 ангстрем структура бактериальной рибосомы.
Предмет
Достижения молекулярной биологии в познании живой природы трудно переоценить. Больших успехов удалось достичь благодаря удачной концепции исследований: сложные биологические процессы рассматриваются с позиции отдельных молекулярных систем, что позволяет применять точные физико-химические методы исследования. Это также привлекло в эту область науки много великих умов из смежных направлений: химии, физики, цитологии, вирусологии , что также благотворно повлияло на масштабы и скорость развития научных знаний в этой области. Столь значимые открытия, как определение структуры ДНК, расшифровка генетического кода, искусственная направленная модификация генома, позволили значительно глубже понять специфику процессов развития организмов и успешно решать многочисленные важнейшие фундаментальные и прикладные научные, медицинские и социальные задачи, которые ещё не так давно считались неразрешимыми.
Предметом изучения молекулярной биологии являются в основном белки, нуклеиновые кислоты и молекулярные комплексы (молекулярные машины) на их основе и процессы, в которых они участвуют.
Нуклеиновые кислоты представляют собой линейные полимеры, состоящие из нуклеотидных звеньев (соединений пятичленного сахара с фосфатной группой при пятом атоме цикла и одного из четырёх азотистых оснований), соединённых между собой сложноэфирной связью фосфатных групп. Таким образом, нуклеиновая кислота - это пентозофосфатный полимер с азотистыми основаниями в качестве боковых заместителей. Химический состав цепочки РНК отличается от ДНК тем, что первая состоит из пятичленного цикла углевода рибозы, тогда как вторая - из дегидрокслилированного производного рибозы - дезоксирибозы. При этом пространственно эти молекулы различаются кардинально, поскольку РНК - это гибкая одноцепочечная молекула, тогда как ДНК - это двуцепочечная молекула.
Белки - это линейные полимеры, представляющие собой цепочки альфа-аминокислот, соединённых между собой пептидной связью, откуда их второе название - полипептиды. В состав природных белков входит множество различных аминокислотных звеньев - у человека до 20 -, что определяет широкое разнообразие функциональных свойств этих молекул. Те или иные белки принимают участие почти в каждом процессе в организме и выполняют множество задач: играют роль клеточного строительного материала, обеспечивают транспорт веществ и ионов, катализируют химические реакции, - список этот очень длинный. Белки образуют устойчивые молекулярные конформации различного уровня организации (вторичные и третичные структуры) и молекулярные комплексы, что ещё больше расширяет их функционал. Эти молекулы могут обладать высокой специфичностью к выполнению каких-либо задач благодаря образованию сложной пространственной глобулярной структуры. Большое разнообразие белков обеспечивает постоянный интерес учёных к этому виду молекул.
Современные представления о предмете молекулярной биологии основаны на обобщении, выдвинутом впервые в 1958 году Фрэнсисом Криком как центральная догма молекулярной биологии. Суть её заключалась в утверждении, что генетическая информация в живых организмах проходит строго определённые этапы реализации: копирование из ДНК в ДНК входе наследования, из ДНК в РНК, а затем из РНК в белок, причём обратный переход не осуществим. Это утверждение было справедливо лишь от части, поэтому впоследствии центральная догма была поправлена с оглядкой на открывшиеся новые данные.
На данный момент известно несколько путей реализации генетического материала, представляющих различные последовательности осуществления трёх видов существования генетической информации: ДНК, РНК и белок. В девяти возможных путях реализации выделяют три группы: это три общих превращения (general), осуществляющиеся в норме в большинстве живых организмов; три особых превращения(special), осуществляющиеся в некоторых вирусах или в особых лабораторных условиях; три неизвестных превращения (unknown), осуществление которых, как считается, невозможно.
К общим превращениям относятся следующие пути реализации генетического кода: ДНК→ДНК (репликация), ДНК→РНК (транскрипция), РНК→белок (трансляция).
Для осуществления передачи наследственных признаков родителям необходимо передать потомкам полноценную молекулу ДНК. Процесс, благодаря которому на основе исходной ДНК может быть синтезирована её точная копия, а следовательно, может быть передан генетический материал, называется репликацией. Он осуществляется специальными белками, которые распутывают молекулу (выпрямляют её участок), расплетают двойную спираль и при помощи ДНК-полимеразы создают точную копию исходной молекулы ДНК.
Для обеспечения жизнедеятельности клетки ей необходимо постоянно обращаться к генетическому коду, заложенному в двойной спирали ДНК. Однако эта молекула слишком велика и неповоротлива для применения её в качестве непосредственного источника генетического материала для непрерывного синтеза белка. Поэтому в ходе реализации информации, заложенной в ДНК, есть посредническая стадия: синтез иРНК, представляющей собой небольшую одноцепочечную молекулу, комплементарную определённому отрезку ДНК, кодирующему некоторый белок. Процесс транскрипции обеспечивается РНК-полимеразой и факторами транскрипции. Полученная молекула затем может быть легко доставлена в отдел клетки, ответственный за синтез белка - рибосому.
После попадания и РНК в рибосому наступает заключительная стадия реализации генетической информации. При этом рибосома считывает с иРНК генетический код триплетами , называющимися кодонами и синтезирует на основе получаемой информации соответствующий белок.
В ходе особых превращений генетический код реализуется по схеме РНК→РНК (репликация), РНК→ДНК (обратная транскрипция), ДНК→белок (прямая трансляция). Репликация такого вида реализуется во многих вирусах, где она осуществляется ферментом РНК-зависимой РНК-полимеразой. Аналогичные ферменты находятся и в клетках эукариот, где они связаны с процессом РНК-глушения (silencing). Обратная транскрипция обнаружена в ретровирусах, где она осуществляется под действием фермента обратной транскриптазы, а также в некоторых случаях в эукариотических клетках, например, при теломерном синтезе. Прямая трансляция осуществляется только в искусственных условиях в изолированной системе вне клетки.
Любой из трех возможных переходов генетической информации из белка в белок, РНК или ДНК считается невозможным. Случай воздействия прионов на белки, в результате которого образуется аналогичный прион, условно можно было бы отнести к виду реализации генетической информации белок→белок. Тем не менее, формально он таковым не является, поскольку не затрагивает аминокислотную последовательность в белке.
Любопытна история возникновения термина «центральная догма». Поскольку слово догма в общем случае означает утверждение, которое не подлежит сомнению, а само слово имеет явный религиозный подтекст, выбор его в качестве описания научного факта не совсем правомерен. По признанию самого Фрэнсиса Крика, это была его ошибка. Он хотел придать выдвигаемой теории большей значимости, выделить её на фоне остальных теорий и гипотез; для чего решил использовать это величественное, по его представлению, слово, не понимая его истинного смысла. Название это, однако, прижилось.
Молекулярная биология сегодня
Бурное развитие молекулярной биологии, постоянный интерес к достижениям в этой области со стороны общества и объективная важность исследований привели к возникновению большого числа крупных научно-исследовательских центров молекулярной биологии по всему миру. Среди крупнейших следует упомянуть следующие: лаборатория молекулярной биологии в Кембридже, Королевский институт в Лондоне - в Великобритании; институты молекулярной биологии в Париже, Марселе и Страсбурге, Пастеровский институт - во Франции; отделы молекулярной биологии в Гарвардском университете и Массачусетском технологическом институте, университете в Беркли, в Калифорнийском технологическом институте, в Рокфеллеровском университете, в институте здравоохранения в Бетесде - в США; институты Макса Планка, университеты в Гёттингене и Мюнхене, Центральный институт молекулярной биологии в Берлине, институты в Йене и Халле - в Германии; Каролинский институт в Стокгольме в Швеции.
В России ведущими центрами в этой области являются Институт молекулярной биологии им. В.А.Энгельгардта РАН, Институт молекулярной генетики РАН, Институт биологии гена РАН, Институт физико-химической биологии им. А. Н. Белозерского МГУ им. М.В.Ломоносова, Институт биохимии им. А.Н.Баха РАН и Институт белка РАН в Пущино.
Сегодня область интересов молекулярных биологов охватывает широкий спектр фундаментальных научных вопросов. По-прежнему ведущую роль занимает изучение структуры нуклеиновых кислот и биосинтеза белка, исследования строения и функций различных внутриклеточных структур, и клеточных поверхностей. Также важными направления исследований являются изучение механизмов рецепции и передачи сигналов, молекулярных механизмов транспорта соединений внутри клетки а также из клетки во внешнюю среду и обратно. Cреди основных направлений научного поиска в области прикладной молекулярной биологии одной из наиболее приоритетных является проблема возникновения и развития опухолей. Также очень важным направлением, изучением которого занимается раздел молекулярной биологии - молекулярная генетика, является изучение молекулярных основ возникновения наследственных заболеваний, и вирусных заболеваний, например, СПИДа, а также разработка способов их предупреждения и, возможно, лечения на генном уровне. Широкое применение нашли открытия и разработки молекулярных биологов в судебной медицине. Настоящая революция в области идентификации личности была сделана в 80-х годах учёными из России, США и Великобритании благодаря разработке и внедрению в повседневную практику метода «геномной дактилоскопии» - установления личности по ДНК. Исследования в этой области не прекращаются и по сей день, современные методы позволяют устанавливать личность с вероятностью ошибки одна миллиардная процента. Уже сейчас идёт активная разработка проекта генетического паспорта, что, как предполагается, позволит сильно снизить уровень преступности.
Методология
Сегодня молекулярная биология располагает обширным арсеналом методов, позволяющих решать самые передовые и самые сложные задачи, стоящие перед учёными.
Одним из самых распространённых методов в молекулярной биологии является гель-электрофорез , который решает задачи разделения смеси макромолекул по размеру или по заряду. Почти всегда после разделения макромолекул в геле применяется блоттинг , метод, позволяющий переносить макромолекулы из геля (сорбировать ) на поверхность мембраны для удобства дальнейшей работы с ними, в частности гибридизации . Гибридизация - формирование гибридной ДНК из двух цепей, имеющих различную природу, - метод, играющий важную роль в фундаментальных исследованиях. Он применяется для определения комплементарных отрезков в разных ДНК (ДНК разных видов), с его помощью происходит поиск новых генов, с его помощью была открыта РНК интерференция, а его принцип лёг в основу геномной дактилоскопии.
Большую роль в современной практике молекулярно-биологических исследований играет метод секвенирования - определения последовательности нуклеотидов в нуклеиновых кислотах и аминокислот в белках.
Современную молекулярную биологию невозможно представить без метода полимеразной цепной реакции (ПЦР). Благодаря этому методу осуществляется увеличение количества (амплификация) копий некоторой последовательности ДНК, чтобы позволяет получить из одной молекулы достаточное количество вещества для дальнейшей работы с ним. Аналогичный результат достигается технологией молекулярного клонирования, в которой требующаяся нуклеотидная последовательность внедряется в ДНК бактерии (живых систем), после чего размножение бактерий приводит к необходимому результату. Этот подход технически значительно сложнее, однако позволяет одновременно получать результат экспрессии исследуемой нуклеотидной последовательности.
Также в молекулярно-биологических исследованиях широко применяются методы ультрацентрифугирование (для разделения макромолекул (больших количеств), клеток, органелл), методы электронной и флуоресцентной микроскопии, спектрофотометрические методы, рентгеноструктурный анализ, авторадиография, и т.п.
Благодаря техническому прогрессу и научным изысканиям в области химии, физики, биологии и информатики современное оборудование позволяет выделять, изучать и изменять отдельные гены и процессы, в которые они вовлечены.
Молекулярная биология пережила период бурного развития собственных методов исследования, которыми теперь отличается от биохимии. К ним, в частности, относятся методы генной инженерии , клонирования , искусственной экспрессии и нокаута генов . Поскольку ДНК является материальным носителем генетической информации, молекулярная биология значительно сблизилась с генетикой , и на стыке образовалась молекулярная генетика , являющаяся одновременно разделом генетики и молекулярной биологии. Так же, как молекулярная биология широко применяет вирусы как инструмент исследования, в вирусологии для решения своих задач используют методы молекулярной биологии. Для анализа генетической информации привлекается вычислительная техника, в связи с чем появились новые направления молекулярной генетики, которые иногда считают особыми дисциплинами: биоинформатика , геномика и протеомика .
История развития
Это основополагающее открытие было подготовлено длительным этапом исследований генетики и биохимии вирусов и бактерий .
В 1928 году Фредерик Гриффит впервые показал, что экстракт убитых нагреванием болезнетворных бактерий может передавать признак патогенности неопасным бактериям. Исследование трансформации бактерий в дальнейшем привело к очистке болезнетворного агента, которым, вопреки ожиданиям, оказался не белок , а нуклеиновая кислота . Сама по себе нуклеиновая кислота не опасна, она лишь переносит гены, определяющие патогенность и другие свойства микроорганизма.
В 50-х годах XX века было показано, что у бактерий существует примитивный половой процесс, они способны обмениваться внехромосомной ДНК, плазмидами . Открытие плазмид, как и трансформации , легло в основу распространённой в молекулярной биологии плазмидной технологии . Ещё одним важным для методологии открытием стало обнаружение в начале XX века вирусов бактерий, бактериофагов . Фаги тоже могут переносить генетический материал из одной бактериальной клетки в другую. Заражение бактерий фагами приводит к изменению состава бактериальной РНК . Если без фагов состав РНК сходен с составом ДНК бактерии, то после заражения РНК становится больше похожа на ДНК бактериофага. Тем самым было установлено, что структура РНК определяется структурой ДНК. В свою очередь, скорость синтеза белка в клетках зависит от количества РНК-белковых комплексов. Так была сформулирована центральная догма молекулярной биологии : ДНК ↔ РНК → белок.
Дальнейшее развитие молекулярной биологии сопровождалось как развитием её методологии, в частности, изобретением метода определения нуклеотидной последовательности ДНК (У. Гилберт и Ф. Сенгер , Нобелевская премия по химии 1980 года), так и новыми открытиями в области исследований строения и функционирования генов (см. История генетики). К началу XXI века были получены данные о первичной структуре всей ДНК человека и целого ряда других организмов, наиболее важных для медицины, сельского хозяйства и научных исследований, что привело к возникновению нескольких новых направлений в биологии: геномики, биоинформатики и др.
См. также
- Молекулярная биология (журнал)
- Транскриптомика
- Молекулярная палеонтология
- EMBO - Европейская организация молекулярных биологов
Литература
- Сингер М., Берг П. Гены и геномы. - Москва, 1998.
- Стент Г., Кэлиндар Р. Молекулярная генетика. - Москва, 1981.
- Sambrook J., Fritsch E.F., Maniatis T. Molecular Cloning. - 1989.
- Патрушев Л. И. Экспрессия генов. - М.: Наука, 2000. - 000 с., ил. ISBN 5-02-001890-2
Ссылки
Wikimedia Foundation . 2010 .
- Ардатовский район Нижегородской области
- Арзамасский район Нижегородской области
Смотреть что такое "Молекулярная биология" в других словарях:
МОЛЕКУЛЯРНАЯ БИОЛОГИЯ - изучает осн. свойства и проявления жизни на молекулярном уровне. Важнейшими направлениями в М. б. являются исследования структурно функциональной организации генетического аппарата клеток и механизма реализации наследственной информации… … Биологический энциклопедический словарь
МОЛЕКУЛЯРНАЯ БИОЛОГИЯ - исследует основные свойства и проявления жизни на молекулярном уровне. Выясняет, каким образом и в какой мере рост и развитие организмов, хранение и передача наследственной информации, превращение энергии в живых клетках и др. явления обусловлены … Большой Энциклопедический словарь
МОЛЕКУЛЯРНАЯ БИОЛОГИЯ Современная энциклопедия
МОЛЕКУЛЯРНАЯ БИОЛОГИЯ - МОЛЕКУЛЯРНАЯ БИОЛОГИЯ, биологическое изучение строения и функционирования МОЛЕКУЛ, из которых состоят живые организмы. К основным сферам изучения относятся физические и химические свойства белков и НУКЛЕИНОВЫХ КИСЛОТ, таких как ДНК. см. также… … Научно-технический энциклопедический словарь
молекулярная биология - раздел биол., который исследует основные свойства и проявления жизни на молекулярном уровне. Выясняет, каким образом и в какой мере рост и развитие организмов, хранение и передача наследственной информации, превращение энергии в живых клетках и… … Словарь микробиологии
молекулярная биология - — Тематики биотехнологии EN molecular biology … Справочник технического переводчика
Молекулярная биология - МОЛЕКУЛЯРНАЯ БИОЛОГИЯ, исследует основные свойства и проявления жизни на молекулярном уровне. Выясняет, каким образом и в какой мере рост и развитие организмов, хранение и передача наследственной информации, превращение энергии в живых клетках и… … Иллюстрированный энциклопедический словарь
Молекулярная биология - наука, ставящая своей задачей познание природы явлений жизнедеятельности путём изучения биологических объектов и систем на уровне, приближающемся к молекулярному, а в ряде случаев и достигающем этого предела. Конечной целью при этом… … Большая советская энциклопедия
МОЛЕКУЛЯРНАЯ БИОЛОГИЯ - изучает явления жизни на уровне макромолекул (гл. обр. белков и нуклеиновых к т) в бесклеточных структурах (рибосомы и др.), в вирусах, а также в клетках. Цель М. б. установление роли и механизма функционирования этих макромолекул на основе… … Химическая энциклопедия
молекулярная биология - исследует основные свойства и проявления жизни на молекулярном уровне. Выясняет, каким образом и в какой мере рост и развитие организмов, хранение и передача наследственной информации, превращение энергии в живых клетках и другие явления… … Энциклопедический словарь
Книги
- Молекулярная биология клетки. Сборник задач , Дж. Уилсон, Т. Хант. Книга американских авторов - приложение ко 2 - му изданию учебника `Молекулярная биология клетки` Б. Албертса, Д. Брея, Дж. Льюиса и др. Содержит вопросы и задачи, цель которых - углубить…
1. Введение.
Предмет, задачи и методы молекулярной биологии и генетики. Значение "классической" генетики и генетики микроорганизмов в становлении молекулярной биологии и генной инженерии. Понятие гена в "классической" и молекулярной генетике, его эволюция. Вклад методологии генной инженерии в развитие молекулярной генетики. Прикладное значение генной инженерии для биотехнологии.
2. Молекулярные основы наследственности.
Понятие о клетке, ее макромолекулярный состав. Природа генетического материала. История доказательства генетической функции ДНК.
2.1. Различные виды нуклеиновых кислот. Биологические функции нуклеиновых кислот. Химическое строение, пространственная структура и физические свойства нуклеиновых кислот. Особенности строения генетического материала про - и эукариот. Комплементарные пары оснований Уотсона-Крика. Генетический код. История расшифровки генетического кода. Основные свойства кода: триплетность, код без запятых, вырожденность. Особенности кодового словаря, семьи кодонов, смысловые и «бессмысленные» кодоны. Кольцевые молекулы ДНК и понятие о сверхспирализации ДНК. Топоизомеры ДНК и их типы. Механизмы действия топоизомераз. ДНК-гираза бактерий.
2.2. Транскрипция ДНК. РНК-полимераза прокариот, ее субъединичная и трехмерная структуры. Разнообразие сигма-факторов. Промотор генов прокариот, его структурные элементы. Стадии транскрипционного цикла. Инициация, образование “открытого комплекса”, элонгация и терминация транскрипции. Аттенюация транскрипции. Регуляция экспрессии триптофанового оперона. “Рибопереключатели”. Механизмы терминации транскрипции. Негативная и позитивная регуляция транскрипции. Лактозный оперон. Регуляция транскрипции в развитии фага лямбда. Принципы узнавания ДНК регуляторными белками (САР-белок и репрессор фага лямбда). Особенности транскрипции у эукариот. Процессинг РНК у эукариот. Кепирование, сплайсинг и полиаденилирование транскриптов. Механизмы сплайсинга. Роль малых ядерных РНК и белковых факторов. Альтернативный сплайсинг, примеры.
2.3. Трансляция , ее этапы, функция рибосом. Локализация рибосом в клетке. Прокариотический и эукариотический типы рибосом; 70S и 80S рибосомы. Морфология рибосом. Подразделение на субчастицы (субъединицы). Кодон-зависимое связывание аминоацил-тРНК в элонгационном цикле. Кодон-антикодоновое взаимодействие. Участие фактора элонгации EF1 (EF-Tu) в связывании аминоацил-тРНК с рибосомой. Фактор элонгации EF1В (EF-Ts), его функция, последовательность реакций с его участием. Антибиотики, воздействующие на этап кодон-зависимого связывания аминоацил-тРНК с рибосомой. Аминогликозидые антибиотики (стрептомицин, неомицин, канамицин, гентамицин и др.), механизм их действия. Тетрациклины как ингибиторы связывания аминоацил-тРНК с рибосомой. Инициация трансляции. Основные этапы процесса инициации. Инициация трансляции у прокариот: факторы инициации, инициаторные кодоны, 3¢-конец РНК малой рибосомной субчастицы и последовательность Шайна-Дальгарно в мРНК. Инициация трансляции у эукариот: факторы инициации, инициаторные кодоны, 5¢-нетранслируемая область и кэп-зависимая «концевая» инициация. «Внутренняя» кэп-независимая инициация у эукариот. Транспептидация. Ингибиторы транспептидации: хлорамфеникол, линкомицин, амицетин, стрептограмины, анизомицин. Транслокация. Участие фактора элонгации EF2 (EF-G) и ГТФ. Ингибиторы транслокации: фусидовая кислота, виомицин, их механизмы действия. Терминация трансляции. Терминирующие кодоны. Белковые факторы терминации прокариот и эукариот; два класса факторов терминации и механизмы их действия. Регуляция трансляции у прокариот.
2.4. Репликация ДНК и ее генетический контроль. Полимеразы, участвующие в репликации, характеристика их ферментативных активностей. Точность воспроизведения ДНК. Роль стерических взаимодействий между парами оснований ДНК при репликации. Полимеразы I, II и III E. coli. Субъединицы полимеразы III. Вилка репликации, “ведущая” и “отстающая” нити при репликации. Фрагменты Оказаки. Комплекс белков в репликационной вилке. Регуляция инициации репликации у E. соli. Терминация репликации у бактерий. Особенности регуляции репликации плазмид. Двунаправленная репликация и репликация по типу катящегося кольца.
2.5. Рекомбинация , ее типы и модели. Общая или гомологичная рекомбинация. Двухнитевые разрывы ДНК, инициирующие рекомбинацию. Роль рекомбинации в пострепликативной репарации двухнитевых разрывов. Структура Холлидея в модели рекомбинации. Энзимология общей рекомбинации у E. coli. RecBCD комплекс. RecA белок. Роль pекомбинации в обеспечении синтеза ДНК при повреждениях ДНК, прерывающих репликацию. Рекомбинация у эукариот. Ферменты рекомбинации у эукариот. Сайт-специфичная рекомбинация. Различия молекулярных механизмов общей и сайт-специфичной рекомбинации. Классификация рекомбиназ. Типы хромосомных перестроек, осуществляемых при сайт-специфичной рекомбинации. Регуляторная роль сайт-специфичной рекомбинации у бактерий. Конструирование хромосом многоклеточных эукариот с помощью системы сайт-специфичной рекомбинации фага.
2.6. Репарация ДНК. Классификация типов репарации. Прямая репарация тиминовых димеров и метилированного гуанина. Вырезание оснований. Гликозилазы. Механизм репарации неспаренных нуклеотидов (mismatch репарация). Выбор репарируемой нити ДНК. SOS-репарация. Свойства ДНК полимераз, участвующих в SOS-репарации у прокариот и эукариот. Представление об “адаптивных мутациях” у бактерий. Репарация двухнитевых разрывов: гомологичная пострепликативная рекомбинация и объединение негомологичных концов молекулы ДНК. Взаимосвязь процессов репликации, рекомбинации и репарации.
3. Мутационный процесс.
Роль биохимических мутантов в формировании теории один ген – один фермент. Классификация мутаций. Точковые мутации и хромосомные перестройки, механизм их образования. Спонтанный и индуцированный мутагенез. Классификация мутагенов. Молекулярный механизм мутагенеза. Взаимосвязь мутагенеза и репарации. Идентификация и селекция мутантов. Супрессия: внутригенная, межгенная и фенотипическая.
4. Внехромосомные генетические элементы.
Плазмиды, их строение и классификация. Половой фактор F, его строение и жизненный цикл. Роль фактора F в мобилизации хромосомного переноса. Образование доноров типа Hfr и F". Механизм конъюгации. Бактериофаги, их структура и жизненный цикл. Вирулентные и умеренные бактериофаги. Лизогения и трансдукция. Общая и специфическая трансдукция. Мигрирующие генетические элементы: транспозоны и IS-последовательности, их роль в генетическом обмене. ДНК-транспозоны в геномах прокариот и эукариот. IS-последовательности бактерий, их структура. IS-последовательности как компонент F-фактора бактерий, определяющего способность передачи генетического материала при конъюгации. Транспозоны бактерий и эукариотических организмов. Прямой нерепликативный и репликативный механизмы транспозиций. Представление о горизонтальном переносе транспозонов и их роли в структурных перерстройках (эктопическая рекомбинация) и в эволюции генома.
5. Исследование структуры и функции гена.
Элементы генетического анализа. Цис-транс комплементационный тест. Генетическое картирование с использованием конъюгации, трансдукции и трансформации. Построение генетических карт. Тонкое генетическое картирование. Физический анализ структуры гена. Гетеродуплексный анализ. Рестрикционный анализ. Методы секвенирования. Полимеразная цепная реакция. Выявление функции гена.
6. Регуляция экспрессии генов. Концепции оперона и регулона. Контроль на уровне инициации транскрипции. Промотор, оператор и регуляторные белки. Позитивный и негативный контроль экспрессии генов. Контроль на уровне терминации транскрипции. Катаболит-контролируемые опероны: модели лактозного, галактозного, арабинозного и мальтозного оперонов. Аттенюатор-контролируемые опероны: модель триптофанового оперона. Мультивалентная регуляция экспрессии генов. Глобальные системы регуляции. Регуляторный ответ на стрессы. Посттранскрипционный контроль. Сигальная трансдукция. Регуляция с участием РНК: малые РНК, сенсорные РНК.
7. Основы генной инженерии. Ферменты рестрикции и модификации. Выделение и клонирование генов. Векторы для молекулярного клонирования. Принципы конструирования рекомбинантных ДНК и их введения в реципиентные клетки. Прикладные аспекты генной инженерии.
а). Основная литература:
1. Уотсон Дж., Туз Дж., Рекомбинантные ДНК: Краткий курс. – М.: Мир, 1986.
2. Гены. – М.: Мир. 1987.
3. Молекулярная биология: структура и биосинтез нуклеиновых кислот. / Под ред. . – М. Высшая шк. 1990.
4. , – Молекулярная биотехнология. М. 2002.
5. Спирин рибосомы и биосинтез белка. – М.: Высшая школа, 1986.
б). Дополнительная литература:
1. Хесин генома. – М.: Наука. 1984.
2. Рыбчин генетической инженерии. – СПб.: СПбГТУ. 1999.
3. Патрушев генов. – М.: Наука, 2000.
4. Современная микробиология. Прокариоты (в 2-х тт.). – М.: Мир, 2005.
5. М. Сингер, П. Берг. Гены и геномы. – М.: Мир, 1998.
6. Щелкунов инженерия. – Новосибирск: Из-во Сиб. Унив., 2004.
7. Степанов биология. Структура и функции белков. – М.: В. Ш., 1996.
(Molekularbiologe/-biologin)
-
Тип
Профессия после диплома
-
Зарплата
3667-5623 € в мес
Молекулярные биологи исследуют молекулярные процессы как основу всех жизненных процессов. На основании полученных результатов они разрабатывают концепции использования биохимических процессов, например, в медицинских исследованиях и диагностике или в биотехнологии. Кроме того, они могут быть вовлечены в производство фармацевтических продуктов, разработку продуктов, обеспечение качества или фармацевтический консалтинг.
Обязанности Молекулярного Биолога
Молекулярные биологи могут работать в разных областях. Например, они касаются использования результатов исследований для производства в таких областях, как генная инженерия, химия белка или фармакология (открытие лекарств). В химической и фармацевтической промышленности они способствуют внедрению недавно разработанных продуктов из исследований в производство, сбыта продукции и консультирования пользователей.
В научных исследованиях молекулярные биологи изучают химико-физические свойства органических соединений, а также химические процессы (в области клеточного метаболизма) в живых организмах и публикуют результаты исследований. В высших учебных заведениях они обучают студентов, готовятся к лекциям и семинарам, проверяют письменную работу и принимают экзамены. Самостоятельная научная деятельность возможна только после получения степени магистра и доктора наук.
Где работают Молекулярные Биологи
Молекулярные биологи находят работу, например
- в научно-исследовательских институтах, например, в областях науки и медицины
- в высших учебных заведениях
- в химико-фармацевтической промышленности
- в департаментах по охране окружающей среды
Зарплата Молекулярного Биолога
Уровень зарплаты, которую получают Молекулярные Биологи в Германии составляет
- от 3667€ до 5623€ в мес
(по данным различных статистических бюро и служб занятости в Германии)
Задачи и обязанности Молекулярного Биолога в подробностях
В чем суть профессии Молекулярный Биолог
Молекулярные биологи исследуют молекулярные процессы как основу всех жизненных процессов. На основании полученных результатов они разрабатывают концепции использования биохимических процессов, например, в медицинских исследованиях и диагностике или в биотехнологии. Кроме того, они могут быть вовлечены в производство фармацевтических продуктов, разработку продуктов, обеспечение качества или фармацевтический консалтинг.
Призвание Молекулярная биология
Молекулярная биология или молекулярная генетика занимается изучением структуры и биосинтеза нуклеиновых кислот и процессов, связанных с передачей и реализацией этой информации в форме белков. Это позволяет понять болезненные нарушения этих функций и, возможно, вылечить их с помощью генной терапии. Существуют интерфейсы для биотехнологии и генной инженерии, в которых создаются простые организмы, такие как бактерии и дрожжи, чтобы сделать вещества, представляющие фармакологический или коммерческий интерес, доступными в промышленном масштабе благодаря целевым мутациям.
Теория и практика Молекулярной Биологии
Химико-фармацевтическая промышленность предлагает многочисленные области занятости для молекулярных биологов. В промышленных условиях они анализируют процессы биотрансформации или разрабатывают и улучшают процессы микробиологического производства активных ингредиентов и фармацевтических промежуточных продуктов. Кроме того, они участвуют в переходе недавно разработанных продуктов из стадии исследований в производство. Выполняя задачи проверки, они гарантируют, что производственные мощности, оборудование, аналитические методы и все этапы производства чувствительных продуктов, таких как фармацевтические препараты, всегда соответствуют требуемым стандартам качества. Кроме того, молекулярные биологи консультируют пользователей по использованию новых продуктов.
Для руководящих должностей часто требуется магистерская программа.
Молекулярные Биологи в Научных исследованиях и Образовании
В области науки и исследований молекулярные биологи занимаются такими темами, как распознавание, транспорт, свертывание и кодификация белков в клетке. Результаты исследований, которые являются основой для практического применения в различных областях, публикуют их и, таким образом, делают их доступными для других ученых и студентов. На конференциях и конгрессах они обсуждают и презентуют результаты научной деятельности. Молекулярные биологи проводят лекции и семинары, руководят научной работой и принимают экзамены.
Самостоятельная научная деятельность требует степени магистра и доктора.
Успехи в изучении нуклеиновых кислот и биосинтеза белка привели к созданию ряда методов, имеющих большое прикладное значение в медицине, сельском хозяйстве и ряде других отраслей.
После того, как был изучен генетический код и основные принципы хранения и реализации наследственной информации, развитие молекулярной биологии зашло в тупик, так как не было методов, которые позволяли манипулировать генами, выделять и изменять их. Появление этих методов произошло в 1970-1980х годах. Это дало мощный толчок развитию этой области науки, которая и сегодня переживает период расцвета. Прежде всего, эти методы касаются получения индивидуальных генов и их введения в клетки других организмов (молекулярное клонирование и трансгенез, ПЦР), а также методов определения последовательности нуклеотидов в генах (секвенирования ДНК и РНК). Ниже эти методы будут рассмотрены более подробно. Мы начнем с простейшего базового метода - электрофореза и затем перейдем к более сложным методам.
ЭЛЕКТРОФОРЕЗ ДНК
Это базовый метод работы с ДНК, применяющийся вместе с практическими всеми другими методами для выделения нужных молекул и анализа результатов. Для разделения фрагментов ДНК по длине применяется метод электрофореза в геле. ДНК - кислота, ее молекулы содержат остатки фосфорной кислоты, которые отщепляют протон и приобретают отрицательный заряд (рис. 1).
Поэтому в электрическом поле молекулы ДНК движутся к аноду - положительно заряженному электроду. Это происходит в растворе электролитов, содержащем ионы-носители заряда, благодаря чему этот раствор проводит ток. Чтобы разделить фрагменты, применяется плотный гель из полимеров (агарозы либо полиакриламида). Молекулы ДНК "запутываются" в нем тем больше, чем они длиннее, и поэтому наиболее длинные молекулы движутся медленнее всего, а наиболее короткие - быстрее всего (рис. 2). Заблаговременно или после электрофореза гель обрабатывают красителями, связывающимися с ДНК и флуоресцирующими в ультрафиолетовом свете, и получают картину полос в геле (см. рис. 3). Для определения длин фрагментов ДНК образца их сравнивают с маркером - набором фрагментов стандартных длин, нанесенных параллельно на тот же гель (рис. 4).



Важнейшими инструментами для работы с ДНК являются ферменты, осуществляющие превращения ДНК в живых клетках: ДНК-полимеразы, ДНК-лигазы и рестрикционные эндонуклеазы, или рестриктазы. ДНК-полимеразы осуществляют матричный синтез ДНК, что позволяет размножать ДНК в пробирке. ДНК-лигазы сшивают между собой молекулы ДНК или залечивают бреши них. Рестрикционные эндонуклеазы , или рестриктазы , разрезают молекулы ДНК по строго определённым последовательностям, что позволяет вырезать отдельные фрагменты из общей массы ДНК. Эти фрагменты могут в каких-то случаях содержать отдельные гены.
рестриктазы
Последовательности, узнаваемые рестриктазами, симметричны, и разрывы могут происходить в середине такой последовательности или со сдвигом (в одном и том же месте в обеих нитях ДНК). Схема действия разных типов рестриктаз показана на рис. 1. В первом случае получаются так называемые «тупые» концы, а во втором – «липкие» концы. В случае «липких» концов дна цепь оказывается короче другой, образуется однонитевой участок с симметричной последовательностью, одинаковой на обоих образующихся концах.
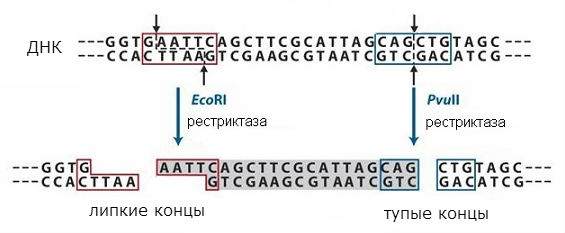
Концевые последовательности будут одинаковыми при расщеплении любой ДНК данной рестриктазой и могут снова соединяться, так как имеют комплементарные последовательности. Их можно сшить с помощью ДНК-лигазы и получить единую молекулу. Таким образом удаётся объединить фрагменты двух разных ДНК и получить так называемые рекомбинантные ДНК . Этот подход используется в методе молекулярного клонирования, позволяющего получить индивидуальные гены и ввести их в клетки, которые могут образовывать закодированный в гене белок.
молекулярное клонирование
В молекулярном клонировании используется две молекулы ДНК - вставка, содержащая интересующий ген, и вектор - ДНК, выступающая в роли носителя. Вставку "вшивают" в вектор пр помощи ферментов, получая новую, рекомбинантную молекулу ДНК, затем эту молекулу внедряют в клетки-хозяева, и эти клетки образуют колонии на питательной среде. Колония - это потомство одной клетки, то есть клон, все клетки колонии генетически идентичны и содержат одну и ту же рекомбинантную ДНК. Отсюда термин "молекулярное клонирование", то есть получение клона клеток, содержащих интересующий нас фрагмент ДНК. После того, как колонии, содержащие интересующую нас вставку, получены, можно различными методами характеризовать эту вставку, например, определить ее точную последовательность. Также клетки могут производить кодируемый вставкой белок, если она содержит функциональный ген.
При внедрении рекомбинантной молекулы в клетки происходит генетическая трансформация этих клеток. Трансформация - процесс поглощения клеткой организма свободной молекулы ДНК из среды и встраивания её в геном, что приводит к появлению у такой клетки новых для неё наследуемых признаков, характерных для организма-донора ДНК. Например, если встраиваемая молекула содержит ген устойчивости к антибиотику ампициллину, то трансформированные бактерии будут расти в его присутствии. До трансформации ампициллин вызывал их гибель, то есть у трансформированных клеток возникает новый признак.
ВЕКТОРЫ
Вектор должен обладать рядом свойств:
Во-первых, это относительно небольшая молекула ДНК, чтобы ей было легко манипулировать.
Во-вторых, для того, чтобы ДНК сохранялась и размножалась в клетке, она должна содержать определённую последовательность, обеспечивающую её репликацию (точку начала репликации, или origin of replication).
В-третьих, она должна содержать ген-маркер , который обеспечивает отбор только тех клеток, в которые попал вектор. Обычно это гены устойчивости к антибиотикам - тогда в присутствии антибиотика все не содержащие вектора клетки погибают.
Клонирование генов чаще всего проводят в клетках бактерий, так как они просты в культивировании и быстро размножаются. В клетке бактерии обычно присутствует одна большая кольцевая молекула ДНК, длиной в несколько миллионов пар нуклеотидов, содержащая все необходимые бактерии гены - бактериальная хромосома. Кроме неё в некоторых бактериях существуют небольшие (несколько тысяч пар нуклеотидов) кольцевые ДНК, называемые плазмидами (рис. 2). Они, также как и основная ДНК, содержат последовательность нуклеотидов, обеспечивающую способность ДНК реплицироваться (ori). Плазмиды реплицируются независимо от основной (хромосомной) ДНК, поэтому присутствуют в клетке в большом количестве копий. Многие из таких плазмид несут гены устойчивости к антибиотикам, что позволяет отличить клетки, несущие плазмиду, от обычных клеток. Чаще используются плазмиды, несущие два гена, обеспечивающие устойчивость к двум антибиотикам, например, к тетрациклину и апмицилину. Существуют простые методы выделения таких плазмидных ДНК, свободных от ДНК основной хромосомы бактерии.

ЗНАЧЕНИЕ ТРАНСГЕНЕЗА
Перенос генов из одного организма в другой называют трансгенезом , а такие модифицированные организмы - трансгенными . Методом переноса генов в клетки микроорганизмов получают рекомбинантные белковые препараты для нужд медицины, в частности, человеческие белки, не вызывающие иммунного отторжения - интерфероны, инсулин и другие белковые гормоны, клеточные факторы роста, а также белки для производства вакцин. В более сложных случаях, когда модификация белков проходит правильно только в клетках эукариот, применяют трансгенные клеточные культуры или трансгенных животных, в частности, скот (прежде всего коз), который выделяет необходимые белки в молоко, или же белки выделяют из их крови. Так получают антитела, факторы свертывания крови и другие белки. Методом трансгенеза получают культурные растения, устойчивые к гербицидам и вредителям и обладающие другими полезными свойствами. При помощью трансгенных микроорганизмов очищают сточные воды и борются с загрязнениями, существуют даже трансгенные микробы, которые могут расщеплять нефть. Помимо этого, трансгенные технологии незаменимы в научных исследованиях - развитие биологии сегодня немыслимо без рутинного применения методов модификации и переноса генов.
технология молекулярного клонирования
вставки
Для получения индивидуального гена из какого-либо организма из него выделяют всю хромосомную ДНК и расщепляют её одной или двумя рестриктазами. Ферменты подбирают так, чтобы они не разрезали интересующий нас ген, а делали разрывы по его краям, а в плазмидной ДНК делали 1 разрыв в одном из генов устойчивости, например, к ампицилину.
Процесс молекулярного клонирования включает следующие этапы:
Разрезание и сшивание - конструирование из вставки и вектора единой рекомбинантной молекулы.
Трансформация - внедрение рекомбинантной молекулы в клетки.
Селекция - отбор клеток, получивших вектор со вставкой.
разрезание и сшивание
Плазмидную ДНК обрабатывают теми же рестриктазами, и она превращается в линейную молекулу, если подобрана такая рестриктаза, которая вносит в плазмиду 1 разрыв. В результате на концах всех образующихся фрагментов ДНК оказываются одни и те же липкие концы. При понижении температуры эти концы соединяются случайным образом, и их сшивают ДНК-лигазой (см. рис. 3).

Получают смесь кольцевых ДНК разного состава: некоторые из них будут содержать определённую последовательность ДНК хромосомной ДНК, соединённую с бактериальной ДНК, другие – соединённые вместе фрагменты хромосомной ДНК, а третьи – восстановленную кольцевую плазмиду или её димер (Рис. 4).

трансформация
Далее этой смесью проводят генетическую трансформацию бактерий, не содержащих плазмиды. Трансформация - процесс поглощения клеткой организма свободной молекулы ДНК из среды и встраивания её в геном, что приводит к появлению у такой клетки новых для неё наследуемых признаков, характерных для организма-донора ДНК. В каждую клетку может проникнуть и размножиться там только одна плазмида. Такие клетки помещают на твёрдую питательную среду, в которой содержится антибиотик тетрациклин. Клетки, в которые не попала плазмида, на этой среде расти не будут, а клетки, несущие плазмиду, образуют колонии, в каждой из которых находятся потомки только одной клетки, т.е. все клетки в колонии несут одну и ту же плазмиду (см. рис. 5).

Селекция
Далее стоит задача выделить только клетки, в которые попал вектор со вставкой, и отличить их от клеток, несущих только вектор без вставки или вовсе не несущих вектора. Этот процесс отбора нужных клеток называется селекцией . Для этого применяют селективные маркеры - обычно гены устойчивости к антибиотикам в составе вектора, и селективные среды , содержащие антибиотики или другие вещества, обеспечивающие селекцию.
В рассматриваемом нами примере клетки из колоний, выросших в присутствии ампицилина, пересевают на две среды: в первой есть ампицилин, а во второй – тетрациклин. Колонии, содержащие только плазмиду, вырастут на обеих средах, а колонии, в плазмидах которых находится встроенная хромосомная ДНК на среде с тетрациклином не вырастут (рис. 5). Среди них специальными методами отбирают те, которые содержат интересующий нас ген, выращивают в достаточных количествах и выделяют плазмидную ДНК. Из неё с помощью тех же рестриктаз, которые использовались при получении рекомбинантной ДНК, вырезают интересующий индивидуальный ген. ДНК этого гена может использоваться для определения последовательности нуклеотидов, введения в какой либо организм для получения новых свойств или синтеза нужного белка. Такой метод выделения генов называется молекулярным клонированием .
ФЛУОРЕСЦЕНТНЫЕ БЕЛКИ
В качестве генов-маркеров при исследованиях эукариотических организмов очень удобно использовать флуоресцентные белки. Ген первого флуоресцентного белка, зеленого флуоресцирующего белка (green fluorescent protein, GFP) был выделен из медузы Aqeuorea victoria и внедрен в различные модельные организмы (см. рис. 6) В 2008 году О. Симомура, М. Чалфи и Р. Тсьен получили Нобелевскую премию за открытие и применение этого белка.

Затем были выделены гены других флуоресцентных белков - красного, синего, желтого. Эти гены были модифицированы искусственно, чтобы получить белки с нужными свойствами. Разнообразие флуоресцентных белков показано на рис. 7, где изображена чашка Петри с бактериями, содержащими гены различных флуоресцентных белков.

применение флуоресцентных белков
Ген флуоресцентного белка можно сшивать с геном любого другого белка, тогда при трансляции будет образовываться единый белок - трансляционно слитый белок, или фьюжн (fusion protein), который флуоресцирует. Таким образом можно изучать, например, локализацию (расположение) любых интересующих белков в клетке, их перемещение. При помощи экспрессии флуоресцентных белков только в определенных типах клеток можно помечать клетки этих типов в многоклеточном организме (см. рис. 8 - мозг мыши, в котором отдельные нейроны имеют разные цвета за счет определенной комбинации генов флуоресцентных белков). Флуоресцентные белки - незаменимый инструмент современной молекулярной биологии.

ПЦР
Еще один метод получения генов называется полимеразной цепной реакцией (ПЦР) . В его основе лежит способность ДНК-полимераз достраивать вторую нить ДНК по комплементарной нити, как это происходит в клетках при репликации ДНК.
Точки начала репликации в этом методе задаются двумя небольшими фрагментами ДНК, называемыми затравками,
или праймерами
. Эти затравки комплементарны концам интересующего гена на двух цепях ДНК. Сначала хромосомную ДНК, из которой надо выделить ген, смешивают с затравками и нагревают до 99 о С. Это приводит к разрыву водородных связей и расхождению нитей ДНК. После этого температуру понижают до50-70 о С (в зависимости от длины и последовательности затравок). В этих условиях затравки присоединяются к комплементарным участкам хромосомной ДНК, образуя правильную двойную спираль (см. рис. 9). После этого добавляют смесь всех четырёх нуклеотидов, нужных для синтеза ДНК, и ДНК-полимеразу. Фермент удлиняет затравки, строя двуспиральную ДНК от места прикрепления затравок, т.е. от концов гена, до конца одноцепочечной хромосомной молекулы.
Если теперь снова нагреть смесь, то хромосомные и вновь синтезированные цепи разойдутся. После охлаждения к ним снова присоединятся затравки, которые берутся в большом избытке (см. рис. 10).

На вновь синтезированных цепях они присоединятся не к тому концу, с которого начинался первый синтез а к противоположному, так как цепи ДНК антипараллельны. Поэтому во втором цикле синтеза на таких цепях достроится только последовательность, соответствующая гену (см. рис. 11).
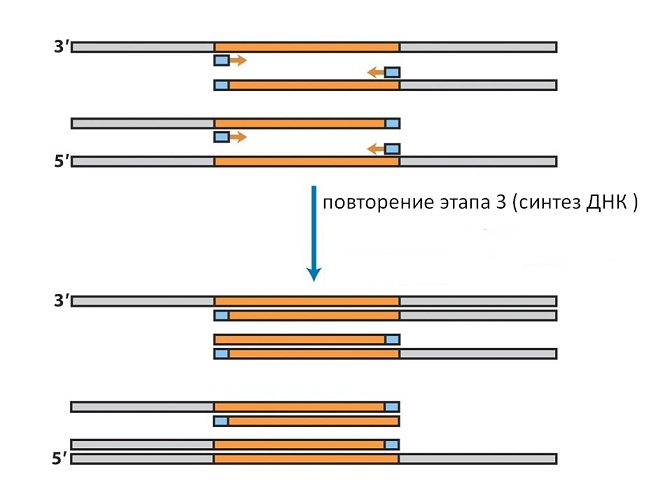
В данном методе применяется ДНК-полимераза из термофильных бактерий, способная выдерживать кипячение и работающая при температурах 70-80 о С, её не надо добавлять каждый раз, а достаточно внести в начале опыта. Повторяя процедуры нагрева и охлаждения в той же последовательности, мы можем в каждом цикле удваивать число последовательностей, ограниченных с двух концов внесёнными затравками (см. рис. 12).

После примерно 25 таких циклов число копий гена увеличится более чем в миллион раз. Такие количества легко можно отделить от внесённой в пробирку хромосомной ДНК и использовать для различных целей.
секвенирование ДНК
Ещё одним важным достижением является разработка методов определения последовательности нуклеотидов в ДНК - секвенирования ДНК (от англ. sequence - последовательность). Для этого необходимо получить чистые от других ДНК гены одним из описанных методов. Затем цепи ДНК разделяют нагреванием и прибавляют к ним затравку, меченую радиоактивным фосфором или флуоресцентной меткой. Обратите внимание, что берётся одна затравка, комплементарная одной цепи. Затем добавляется ДНК полимераза и смесь из 4-х нуклеотидов. Такая смесь делится на 4 части и к каждой добавляется один из нуклеотидов, модифицированный так, что у третьего атома дезоксирибозы он не содержит гидроксильной группы. Если такой нуклеотид включится в синтезируемую цепь ДНК, то её удлинение не сможет продолжаться, т.к. полимеразе некуда будет присоединять следующий нуклеотид. Поэтому синтез ДНК после включения такого нуклеотида обрывается. Таких нуклеотидов, называемых дидезоксинуклеотиды, добавляется значительно меньше, чем обычных, поэтому обрыв цепи происходит лишь изредка и в каждой цепи в разных местах. В результате получается смесь цепей разной длины, на конце каждой из них стоит один и тот же нуклеотид. Таким образом длина цепи соответствует номеру нуклеотида в изучаемой последовательности, например, если у нас был адениловый дидезоксинуклеотид, а полученные цепи имели длину 2, 7 и 12 нуклеотидов, значит в гене во второй, седьмой и двенадцатой позиции стоял аденин. Полученную смесь цепей легко разделить по размерам при помощи электрофореза, а синтезированные цепи выявить по радиоактивности на рентгеновской плёнке (см. рис. 10).

Получается картина, приведённая внизу рисунка, называемая радиоавтографом. Двигаясь по нему снизу вверх и читая буква над колонками каждой зоны мы получим последовательность нуклеотидов, приведённую на рисунке справа от автографа. Оказалось, что синтез останавливается не толоко дидезоксинуклеотидами, но и нуклеотидами, у которых в третьем положении сахара присоединяется какая-нибудь химическая группа, например флюоресцентный краситель. Если каждый нуклеотид пометить своим красителем, то зоны, получаемые при разделении синтезированных цепей, будут светиться разным светом. Это позволяет проводить реакцию в одной пробирке одновременно для всех нуклеотидов и разделяя полученные цепи по длине, идентифицировать нуклеотиды по цвету (см. рис. 11).

Такие методы позволили определить последовательности не только отдельных генов, но и прочитать целые геномы. В настоящее время разработаны ещё более быстрые методы определения последовательностей нуклеотидов в генах. Если парвый геном человека был расшифрован большим международным консорциумом с использованием первого приведённого метода за 12 лет, второй, с использованием второго, за три года, то сейчас это может быть сделано за месяц. Это позволяет предсказывать предрасположенность человека к многим заболеваниям и заранее принимать меры, чтобы избежать их.

")
 Расписание поста рамадан в году татарстан")


